Aspects of the Evolutionary Biology of Pennatulacean Octocorals
Gary C. Williams
Figure 2 Comparative Morphology
Table of ContentsAbstract
Introduction
Historical Review
Phylogenetic Assessment
Ediacaran Frond-like Fossils - A Reassessment
Literature Cited
ABSTRACT
A preliminary assessment of the phylogeny of sea pens is presented, as well as a synopsis of the history of the literature pertaining to the evolution and phylogeny of the Pennatulacea, and a reassessment of the Ediacaran frond-like fossils in light of phylogenetic and fossil evidence. Examination of recently acquired material of broad geographic scope, in addition to older collections and the literature, has allowed for a phylogenetic re-assessment of the group using cladistic analyses.
The published history of evolutionary thought regarding the pennatulaceans spans the period 1870-1916. It was not until 1994 that the subject was resurrected.
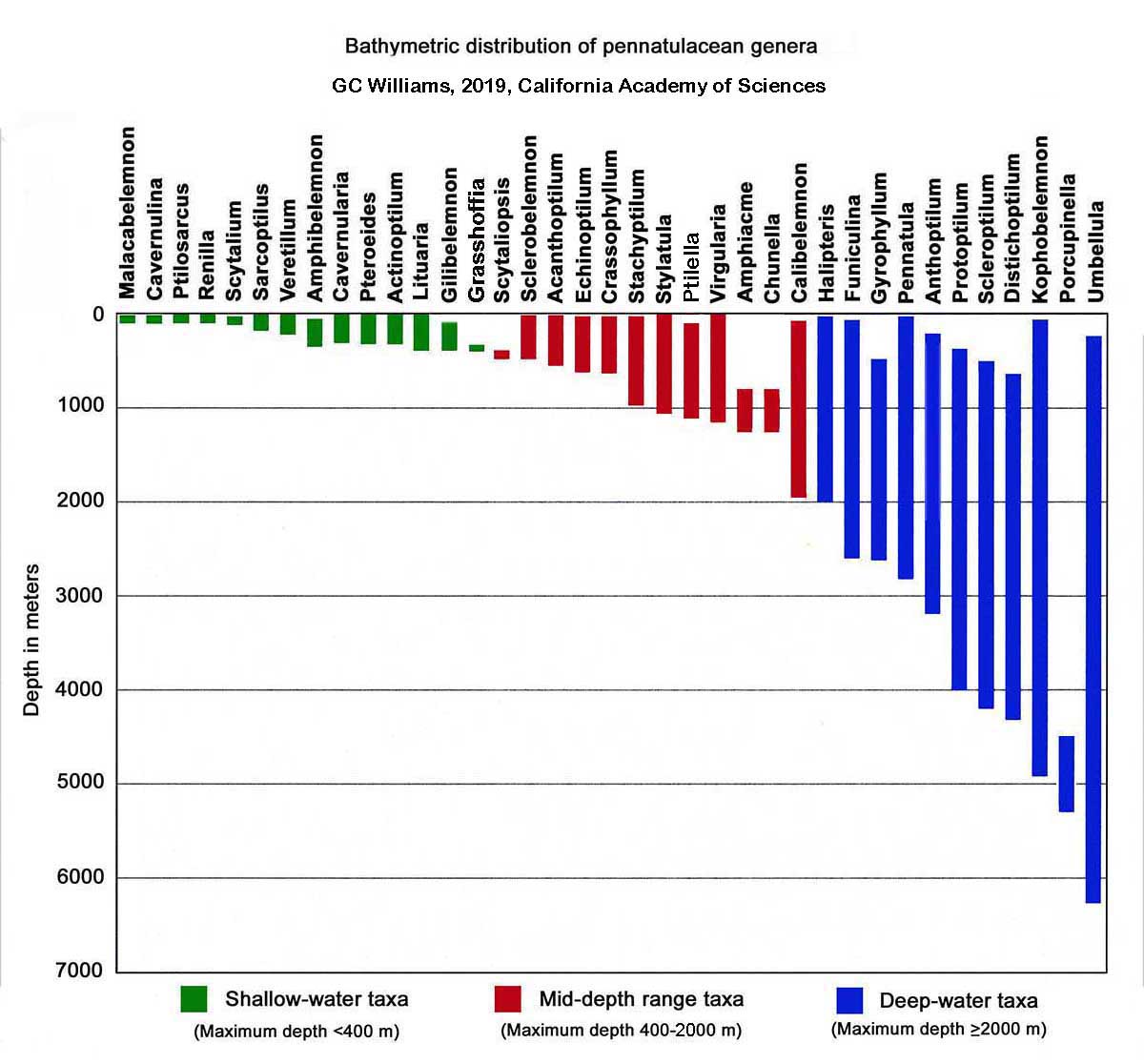
Distributional and phylogenetic data support the hypothesis that the sea pens first differentiated in the shallow-water tropics and then subsequently dispersed and diversified in temperate and polar regions, and to all ocean depths as well as the shallow-water tropics. Primitive shallow-water tropical taxa are represented by Cavernularia and Veretillum - while variously derived, deeper water taxa of widespread distribution include Funiculina, Chunella, Umbellula, Pennatula, Gyrophyllum, Distichoptilum, and Kophobelemnon.
The veretillid sea pens are postulated to represent the most primitive or basal group for the Pennatulacea and at the same time represent a monophyletic clade distinguished from other octocorals by a distinctive set of apomorphies.
The origin of the Pennatulacea is sought in alcyoniid-like ancestors. At least superficial similarities are found between veretillids such as Lituaria and soft coral taxa such as Verseveldtia and Bathyalcyon. If strong evidence is found for alcyoniid ancestors of the pennatulaceans, then the Alcyonacea sensu lato must necessarily be considered paraphyletic.
Morphological and phylogenetic evidence does not support the premise that the Vendian frond-like fossil taxa of the Ediacaran and Burgess shale faunas such as Charniodiscus, Vaizitsinia, and Khatyspytia, and Thaumaptilon represent pennatulacean octocorals. The lateral branches of the frond-like fossils and the polyp leaves of leafy sea pens not only appear to be nonhomologous but not even functionally convergent.
INTRODUCTION
Octocoral systematics at present is often an equivocal and disputatious field (Weinberg, 1976). Our knowledge concerning this group of anthozoan coelenterates is far from adequate (Bayer, 1981). Substantial confusion has arisen in the literature because of a general lack of knowledge concerning variability - some octocoral species can be exceedingly variable and many authors have described separate taxa based on intrapecific variants. Comprehensive collecting in many different geographical localities and detailed comparison of material is essential to adequately assess the degree of phenotypic variation in many taxa due to genetic, geographicical, or ecological differences. In addition, the state of the literature for many taxa is inadequate. Many descriptions are very poor in lack of detail, illustrations, or designation of type or voucher material. Entire groups need comprehensive revision before positive identifications of many taxa are possible (i.e. the gorgonian families Melithaeidae and Ellisellidae).
The Pennatulacea as a group were last monographed by Kükenthal (1915), in which only brief, sparsely-illustrated descriptions were included. Hickson (1916) and Williams (1990a) provided regional faunas in which many taxa were included. Williams (1989) reviewed the genus Cavernularia, Williams (1995a) provided a synopsis of the living genera of sea pens, and Williams (1995b) revised the genus Sarcoptilus.
The pennatulaceans or sea pens, are a very distinct and highly specialized group of benthic and sessile coelenterates, found throughout the world's seas at all depths (Hickson, 1909; Bayer, 1956; Williams, 1990). Of the 436 nominal species names appearing in the literature, approximately one half are at present considered valid (Williams, 1995a). Thirty-two genera in fifteen families are currently recognized. The greatest diversity of sea pen taxa is found in the Indo-Pacific. Several pennatulacean species exhibit near-cosmopolitan distributions. The sparse pennatulacaen fossil record extends back to the Cretaceous and Tertiary Periods - represented by Virgularia, Graphularia, and Pteroeides, with likely alcyonacean sclerites from the Silurian (Bengtson, 1981) and controversial and problematic taxa from the Ediacara fauna (Bayer, 1956). Unlike other octocorals, sea pens are characterized by having the bulk of the body composed of a single large primary polyp - the oozooid. Lateral budding of the oozooid body wall gives rise to two or three kinds of secondary polyps (always autozooids and siphonozooids, and rarely mesozooids). The oozooid has a basal fleshy peduncle that acts to anchor the animal in soft substrata such as sand, mud, rubble, or abyssal ooze deposits, by peristaltic contractions and hydrostatic pressure. The secondary polyps are variously arranged on the upper portion of the animal - the rachis - which projects above the surface of the sea bottom.
Pennatulaceans are morphologically a very diverse group that exhibit trends toward bilateral symmetry, lateral processes in the polyp leaves or ridges, localization and concentration of polyps, and reduction in size and number of sclerites.
HISTORICAL REVIEW
A few authors have attempted to deal with the subject of the evolution and phylogenetics of veretillids and other sea pens, these works being antiquated to say the least (1870-1916)! The only modern phylogenetic analysis are those of Williams (1994 & 1995c). Kölliker (1870 [1872]: 449) was the first to address the phylogenetic development of sea pens, where he considered Umbellula along with Protoptilum to be primitive offshoots of the pennatulacean prototype and the veretillids as highly derived forms from Kophobelemnoid ancesters. Koch (1878) countered the second part of Kölliker's premise, viewing the veretillids as transitional forms between the alcyonaceans and the pennatulaceans, "Ich betrachte sie (Pennatuliden) als Verwandte der Alcyoniden, zu denen einige Veretilliden ganz gute Uebergangsformen bilden".
Kölliker (1880: 39) believed the deep sea pennatulaceans to be the primitive forms and probable relicts of an ancient and extinct fauna, "The simpler forms of the Pennatulida, especially those with sessile polyps, inhabit great depths...These simpler forms are probably also the oldest, and may be regarded as the last remnants of an extinct primary creation. The Protoptilidae and the Umbellulidae are the principal representatives of these old forms". Kölliker's argument should be viewed from the perspective of the mid-nineteenth century school of thought that considered the virtually unknown abyss as the home of the primordial ooze - the "Bathybius" of English science and the "Erstschleim" of German science. Although contrary to our modern interpretations, Kölliker's views can be considered discerning in light of the status of knowledge at the time. Kölliker viewed all forms with sessile polyps as primitive and those with polyps emanating from accessory appendages as derived. Our modern interpretation views this character state as only one in a suite of characters that are important in determining relationships and in differentiating primitive versus derived forms. Some of the forms with sessile polyps such as Chunella and Umbellula are now considered highly derived (Fig. 1 & Williams, 1994). In addition, the shallow water veretillids (currently regarded as the most primitive group), were not well known during Kölliker's time. Even with this limitation, however, Koch was perceptive enough to recognize their relevance to the evolution of the sea pens as a whole.
Marshall (1887: 148) disputed Kölliker's 1880 claim (as well as the first part of his 1872 premise) by acknowledging the striking diversity of deep sea forms together with the markedly derived nature of Umbellula, "The most noteworthy point is the great abundance and variety of specimens dredged...from a depth of 555 fathoms...These results...do not lend any material support to Kölliker's conclusion, that 'the simpler forms of Pennatulida, especially those with sessile polyps, inhabit great depths'. Kolliker ranks among primitive forms of Pennatulida the Umbellulidae, which are an essentially deep water family...and cites this distribution in evidence of the view that the lower forms of Pennatulida are, as a rule, deep water forms. Umbellula appears to me, however, to be not a primitive form but a highly modified one...In all these respects Umbellula is far less primitive than Funiculina, which is essentially a shallow water form, attaining its maximum of development at about 30 fathoms depth".
Bourne (1900: 33) considered bilateralism and polyp leaves as derived characters, "The higher members of the Pennatulacea have a distinct bilateral symmetry, due to the zooids being borne like the barbs of a feather on two sides of the rachis only, leaving a sterile band on the two remaining sides". However, he considered the radial symmetry of the Veretillidae and the strongly bilateral symmetry of the Funiculinidae to be separate derivatives of a Protocaulon-like archetype, "The existing families of the Pennatulaceae appear to have diverged from an ancestral form resembling Protocaulon molle". Protocaulon molle Kölliker, 1880, exemplifies bilateral symmetry, and was subsequently considered as an unidentifiable species of the genus Virgularia by Kükenthal (1915: 71). Protoptilum and Funiculina were considered to be members of closely related families by Kükenthal & Broch (1911: 456), and the present study agrees with this premise. In addition, Bourne viewed the Umbellulidae and the various taxa with polyp leaves as two separately derived lineages of the Funiculinidae, while he considered the Veretillidae to be a terminal offshoot of Protocaulon.
Kükenthal & Broch (1911: 556) in their concluding chapter entitled, "Die Stammesgeschichte der Seefedern", gave a detailed account of pennatulacean phylogeny. They supported Marshall's contention that Umbellula is a derived rather than primitive taxon and corroborated Koch's position that the veretillids are the most primitive surviving pennatulaceans. The acceptance of the veretillid genera as primitive sea pens was reiterated by Kükenthal (1912: 563) and Niedermeyer (1913: 263). Kükenthal (1915) produced the first comprehensive taxonomic synthesis of the Pennatulacea, and placed the Veretillidae first in a systematic series for the order.
Hickson (1916: 25) gave a detailed assessement of the evolution of the Pennatulacea. He recognized the dilemma of Bourne's view in the great morphological disparity between an alcyonacean ancestor and Bourne's primitive sea pens, "Bourne...was of opinion that 'the existing families of Pennatulacea appear to have diverged from an ancestral form resembling Protocaulon molle', but the difficulty of this view is that it leaves such a great gap in structure between the Pennatulacea and the other Alcyonaria". Although Hickson viewed the Veretillidae as the most primitive existing family, he disagreed with Kükenthal's 1912 contention that Lituaria is the most primitive genus simply because of the presence of tuberculated sclerites, "Kukenthal...regards the genus Lituaria as the most primitive of all the Pennatulacea and places it at the root of his family tree. With that conclusion I am not entirely in agreement". Hickson viewed the presence of an axis and the occasional hint of bilateral symmetry as ample evidence to regard Lituaria as a somewhat specialized veretillid. Hickson recognized the superficial similarity between the soft coral Verseveldtia trochiforme (Hickson, 1900) (see Williams, 1990b) and various veretillids, and chose it to illustrate the prototype of the Pennatulacea, "...we have an Alcyonacean which shows certain features of resemblance to a Veretillid. It is dimmorphic, the colony shows a marked division into regions corresponding to the rachis and stalk, it is radially symmetrical and it has a very narrow base of attachment. These points of resemblance may be due to convergence but the species gives us a representation of what the hypothetical ancestor may have been like before it became detached and took to an independent existence".
PHYLOGENETIC ASSESSMENT
Williams (1994 & 1995c) represents the first modern phylogenetic analyses regarding the pennatulacaean octocorals. Very few other cladistic works have appeared that pertain to "lower invertebrate" groups. These include Weerdt (1985), Hooper (1987 and 1990) for the demospongian genera Acarnus and Rhabderemia (Porifera), Hoeksema (1989 and 1991) for fungiid scleractinians (Coelenterata: Anthozoa), Cairns (1984a, 1984b, 1987) for stylasterine and scleractinian corals (Coelenterata: Hydrozoa and Anthozoa), Gerhart (1983) for gorgonian octocorals (Coelenterata: Anthozoa), and Schmidt (1972 and 1974) for the anthozoan coelenterates as a whole.
Williams (1994) provided several reasons why cladistic analyses for lower metazoans such as pennatulaceans are often seemingly intractable, explaining the consequent paucity of cladistic work in the literature regarding these taxa. These are (1) the scarcity of good characters and the difficulty of character analysis; (2) a poorly represented and problematic fossil record, which makes identification and comparison with extinct groups difficult or impossible (Conway-Morris, 1991); (3) difficulty of obtaining fresh material of many taxa for study (especially for application of molecular and genetic techniques); (4) a high frequency of homoplasy (parallelisms, convergences, and reversals); and (5) unusually high degrees of intraspecific variability. Solé-Cava & Thorpe (1991: 69) have shown that mean heterozygosity values for sponges and coelenterates are very high compared to other groups of organisms such as vertebrates, arthropods, and plants.
If we accept Hickson's premise that the veretillids are intermediate between the alcyonaceans and the other pennatulaceans, then a detailed study of the veretillids is essential to shed light on the phylogenetic relationships of the pennatulaceans in general as well as the nature of the pennatulacean ancestor and sister group relationships. Such a study of the veretillids is crucial to our understanding of the origin of the Pennatulacea, as opposed to studying a more derived clade such as the pteroeidids, which may be easier to define as a monophyletic group, but would not help define the sea pen ancestor or elucidate broader aspects of pennatulacean phylogeny.
Along with the echinoptilids, the veretillids are considered the least derived of the extant sea pens, and exhibit the greatest diversity in the relatively shallow waters of the Indo-Pacific, while a great variety of more derived forms are present worldwide with unrestricted bathymetric range. The veretillid taxa represent a particularly important group for study since they occupy a basal position in the Pennatulacea, and are therefore ideal candidates for study to help elucidate the nature of the sea pen ancestor and the phylogenetic affinities of the group in general. Highly derived forms such as Virgularia and Pteroeides are also present in the shallow-water tropics and are sympatric with the more primitive taxa, while other highly derived forms such as Chunella and Ombellula are restricted to deep water. It has therefore been postulated by Williams (1994: 734) that the sea pens as a group initially differentiated in the shallow waters of tropical oceans and have subsequently diversified and dispersed to all depths of the temperate and polar regions, as well as the tropics (Figure 1 & 2).
The veretillid genera Cavernularia, Cavernulina, Lituaria, and Veretillum are restricted to shallow-water (0-300 meters in depth but mostly in less than 100 meters), and are distributed from southern Europe and western Africa, throughout much of the Indo-Pacific, to parts of the Panamic Province (Mexico to Ecuador). The greatest species diversity occurs in the western Pacific triangle formed by the Philippines, Indonesia, and New Guinea (Williams, 1994: 732). Of the 61 nominal species names appearing in the literature, at least 35 are probably valid (Table 1). Williams (1995a: 103) expresses doubt concerning the validity of the genus Cavernulina and its differentiation from Cavernularia. At present, the distinctions between the various species of veretillids rely mainly on the differences in size and shape of the sclerites (spicules) and development of the axis. The axis varies from being well-developed as in some species of Lituaria to rudimentary or absent althogether in some species of Cavernularia. New taxa are anticipated to be discovered and described as new collecting and examination of previously collected material is undertaken.
The veretillids are a group of pennatuacean octocorals that are very unlike other sea pens. They are thought to occupy a basal position in the phylogeny of the pennatulaceans, and at the same time represent a monophyletic group. Because of this, they no doubt have symplesiomorphic characters which do not serve to segregate them as a monophyletic clade. However, preliminary investigation has revealed several apomorphies to define the veretillids as a distinguishable and unique clade of pennatulaceans - a peduncle, a primary polyp, radially arranged and evenly distributed polyps around the entire rachis, smooth sclerites, and fully retractile polyps.
The sister group to the veretillids are hypothesized to be the echinoptilids (figure 1), which share radial symmetry with the veretillids, but have three-flanged sclerites and permanent polyp calyces unlike the veretillids, which have smooth sclerites and completely retractile polyps. The clade containing the veretillids and the echinoptilids thus represent a clade with basal position for the Pennatulacaea. The nature of the symplesiomorphy of such a group as the veretillids, can only be elucidated with detailed comparison of suspected alcyoniid outgroup taxa such as Bathalcyon and related pennatulacean taxa such as the echinoptilids (Actinoptilum and Echinoptilum) and the kophobelemnids (which include Sclerobelemnon).
Based strictly on morphogical character comparison, Hickson (1916: 26) and Williams (1994: 733) hypothesized regarding the origin of the pennatulaceans and the nature of the pennatulacean ancestor. Various taxa of the large and diverse soft coral family Alcyoniidae share more characters with the veretillids than any other alcyonacean group. Similarities regarding dimorphism, radial symmetry, clearly defined polyparia, and capstan-like sclerites are shared by certain veretillids (Lituaria) and alyconiid taxa such as Verseveldita trochiforme (Hickson, 1900) and Verseveldtia bucciniforme Williams, 1990. A large primary polyp with numerous smaller secondary polyps is found in both veretillids and the alcyoniid Bathyalcyon robustum Versluys, 1906. If we accept an alcyoniid ancestor for the pennatulaceans, then the paraphyly of the Alcyonacea (including stolonferans, soft corals, and gorgonians) is implicit.
Table 1 and Figure 1 represent a data matrix and preliminary cladogram to establish the veretillids as a monophyletic group, and to portray the echinoptilids as the sister group of the veretillids, as well as to show the basal position of the veretillids and their relationship to other pennatulaceans. Table 1 shows the character sets that have been established at the present time. Further detailed investigations of many taxa will potentially yield character sets of greater range and relevancy. Characters were polarized using an alcyoniid ancestor (such as Alcyonium) as an outgroup. PAUP (Phylogenetic Analysis Using Parsimony) version 3.1 was used to produce twelve most parsimonious cladograms, each having a CI (Consistency Index) of 0.783. Every tree showed basal positions for the veretillids and echinoptilids, and each as monophyletic groups. The twelve trees showed minor variations in the relative positions of the other seven taxa. All veretillid taxa that have thus far been examined via the literature or collected specimens, represent a monophyletic clade distinguished from other octocorals by the following combination of synapomorphies - a peduncle, radially arranged and evenly distributed polyps around the entire rachis, smooth sclerites, and fully retractile polyps.
Current Reassessment: With the development and increasing sophistication of techniques in molecular phylogenetics over the past two decades, it has been shown that the veretillids are not basal pennatulaceans, but on the contrary are impbedded within the overall pennatulacean topography; that pennatulaceans and ellisellid gorgonians are sister taxa; and that the genus Sclerobelemnon can be aligned with the veretillids, rather than with other kophobelemnids (McFadden, et al. 2006; Chen 2011; Kushida & Reimer 2018).EDIACARAN FROND-LIKE FOSSILS - A REASSESSMENT
An exceptionally rich literature (e.g. Bergström, 1991; Buss & Seilacher, 1994; Conway Morris, 1991; Glaessner, 1979 & 1984; Glaessner & Wade, 1966; Retallack, 1994; and Seilacher, 1989), has arisen to describe and assess the taxonomic status of various pennatulacean-like Precambrian fossils from the Ediacaran fauna, first described from Australia, but subsequently found in Namibia, Russia, and England. These fossils have been defined in a variety ways including - as a phylum of extinct metazoans (Vendobionta), as a separate nonmetazoan kingdom (Vendobionta), as lichens, and as sea pens. These forms are generally either frond-like with an elongate or a bulbous holdfast, and include the genera Charnia, Chariodiscus, Glaessnerina, Vaizitsinia, and Khatyspytia, or are bag-like foliate forms with similarly bulbous holdfasts such as Pteridinium, and Rangea. An additional taxon, very similar to several Ediacaran forms, has recently been described from the Cambrian Burgess Shale of British Columbia - Thaumaptilon walcotti Conway Morris, 1993.
Of the thirty-two genera of extant pennatulaceans, only six have the large, well-developed polyp leaves that superficially resemble those of the ediacaran forms. These six taxa - Pennatula, Ptilosarcus, Gyrophyllum, Sarcoptilus, Crassophyllum, and Pteroeides - are considered the most highly derived pennatulacean taxa (Figure 2, Williams, 1994 & 1995c). Material representing these taxa are not known from any geological period prior to the Tertiary. The phylogenetics of the above six taxa is treated in detail in Williams (1995c).
The Ediacaran forms are for the most part foliate (leaf-like) and have a continuous margin around the leaf-like frond (Fig. 3D). The extant sea pens with polyp leaves, on the other hand, are pennate (feather-like), having numerous lateral appendages attached only at their bases to the central rachis. The lateral appendages of all of these extant taxa are actually separate polyp leaves in two opposite rows along the rachis, containing the polyps. The single frond of the Ediacaran forms cannot be considered homologous or even functionnally convergent with the polyps leaves of extant sea pens. In order for extinct forms such as Thaumaptilon (as depicted by Conway Morris, 1993: 602) to be convergent with extant leafy sea pens that have an erect rather than prostrate habit, the areas between the lateral branches would have to represent slits or elongated openings that perforate completely through the frond, or the "zooids" would have to represnt holes or small perforations through the frond that would allow for water to flow through the organism like a sieve. If we interpret the frond-like fossils as "sieve organisms", then they may have been functionally convergent with the extant leafy sea pens, with an erect rather than prostrate habit, and the perforated frond would then be considered analogous to but not homologous to the rachis and leaves of leafy pennatulaceans.
The separate polyp leaves of sea pens allow water to flow through the upper part of animal in quite a different way, thus allowing for feeding on planktonic organisms utilizing bottom currents. A rachis composed of a single flattened non-perforated plate would undoubtedly produce a high amount of resistance in a strong bottom current. This resistance is greatly reduced in the feather-like sea pens with free and separate leaves that are attached only at the base (Figure 3C). The single frond of the Ediacaran forms appears to be morphologically more like that of the foliate sea pansy Renilla (Fig. 3B), which does not have repeated polyp leaves, and lies flat on the surface of the sea bottom with the peduncle buried in the sediment. The Ediacaran forms may have had a similar life habit, or could have lived totally immersed in the sediment as postulated by Buss & Seilacher (1994: 3).
As previously stated, the least derived extant taxa, including the most primitive sea pens, the veretillids, are not feather-like but rather are vermiform or clavate to cylindrical in shape, lateral appendages resembling polyp leaves are absent altogether (Fig. 3A). None of these taxa are represented in the fossil record. Graphularia and Virgularia, sea pens with relatively weakly developed polyp leaves, are the only other pennatulacean genera represented by fossils, and these are not recorded from strata earlier than the Jurassic.
Based on the paleontological and phylogenetic evidence, as well as that of comparative morphology, it is here hypothesized that the Ediacaran/Burgess Shale "sea pens" such as Thaumaptilon are not fossilized pennatulacaean octocoral coelenterates, but rather represent another unrelated but superficially similar lineage, one without living representatives - a lineage that was probably extinct by the end of the Cambrian. At present, there is no evidence to suggest that the pennatulaceans were present prior to the Mesozoic.
The problematic nature of interpreting the nature of the Ediacaran fossils is in part due, I believe, to the specialized state of science in which workers in different fields concerned with similar issues are unaware of each other's work and findings. It would be more productive if neontologists (i.e., systematists) and Precambrian paleontologists worked together to decipher the affinities of the organisms in the Ediacara and Burgess Shale faunas.
LITERATURE CITED
BAYER, F.M. 1956. Octocorallia. In: Moore, R.C. (ed.), Treatise on invertebrate paleontology, Part F, Coelenterata: 166-231. Geological Society of America and University of Kansas Press, Lawrence.
BAYER, F.M. 1981. Status of knowledge of octocorals of world seas. Seminarios de Biologia Marinha, Academia Brasileira de Ciencias Rio de Janeiro 1981: 3-102.
BENGTSON, S. 1981. Atractosella, a Silurian alcyonacean octocoral. Journal of Paleontology 55(2): 281-294.
BERGSTRÖM, J. 1991. Metazoan evolution around the Precambrian-Cambrian transition. In: Simonetta, A.M. & Conway-Morris, S. (eds.) The early evolution of Metazoa and the significance of problematic taxa. Proceedings of an International Symposium held at the University of Camerino 27-31 March 1989. New York: Cambridge University Press: 25-34.
BOURNE, G.C. 1900. The Anthozoa. In: Lankester, E.R. (ed.), A treatise on zoology, Part 2. 80 pp.
BROCH, H. 1958. Octocorals. Part I. Pennatularians. Discovery Reports 29: 245-280.
BUSS, L.W. & SEILACHER, A. 1994. The Phylum Vendobionta: a sister group of the Eumetazoa? Paleobiology 20(1): 1-4.
CAIRNS, S.D. 1984a. An application of phylogenetic analysis to the Scleractinia: family Fungiidae. Palaeontographica Americana 54: 49-57.
CAIRNS, S.D. 1984b. A generic revision of the Stylasteridae (Coelenterata: Hydrozoa). Part 2, Phylogenetic analysis. Bulletin of Marine Science 35: 38-53.
CAIRNS, S.D. 1987. Evolutionary trends in the Stylasteridae (Cnidaria, Hydrozoa). In: Bouillon, J., Boero, F., Cicogna, F., Cornelius, P.F.S. (eds.), Modern trends in the systematics, ecology, and evolution of hydroids and hydromedusae (Chapter 23, pp. 257-274).
CONWAY MORRIS, S. 1991. Problematic taxa: a problem for biology or biologists? In: Simonetta, A.M. & Conway-Morris, S. (eds.) The early evolution of Metazoa and the significance of problematic taxa. Proceedings of an International Symposium held at the University of Camerino 27-31 March 1989. New York: Cambridge University Press: 19-24.
CONWAY MORRIS, S. 1993. Ediacaran-like fossils in Cambrian Burgess shale-type faunas of North America. Palaeontology 36(3): 593-635.
GLAESSNER, M.F. 1979. Precambrain biogeography and biostratigraphy. Treatise on Invertebrate Paleontology. Pat A Introduction, A79-A118,
GLAESSNER, M.F. 1984. The dawn of animal life - a biohistorical study. Cambridge University Press, Cambridge, 244 pp.
GLAESSNER, M.F. & WADE, M. 1966. The Late Precambrian fossils from Ediacara, South Australia. Palaeontology 9(4): 599-628.
GERHART, D.J. 1983. The chemical systematics of colonial marine animals: an estimated phylogeny of the order Gorgonacea based on terpenoid characters. Biological Bulletin 164: 71-81.
HICKSON, S.J. 1900. The Alcyonaria and Hydrocorallinae of the Cape of Good Hope. Marine Investigations in South Africa 1: 67-96.
HICKSON, S.J. 1909. Coelenterata: Anthozoa. In: Harmer, S.F., Shipley, A.E. (eds.), The Cambridge Natural History 1: 326-411. Macmillan & Co., London.
HICKSON, S.J. 1916. The Pennatulacea of the Siboga Expedition, with a general survey of the order. Siboga Expeditie 14: 1-265.
HOEKSEMA, B. 1989. Taxonomy, phylogeny and biogeography of mushroom corals (Scleractinia: Fungiidae). Zoologische Verhandelingen 254: 1-295.
HOEKSEMA, B. 1991. Evolution of body size in mushroon corals (Scleractinia: Fungiidae) and its ecomorphological consequences. Netherlands Journal of Zoology 41: 112-129.
HOOPER, J.N.A. 1987. New records of Acarnus Gray (Porifera: Demospongiae: Poecilosclerida) from Australia, with a synopsis of the genus. Memoirs of the Queensland Museum 25: 71-105.
HOOPER, J.N.A. 1990. A new species of Rhabderemia Topsent (Porifera: Demospongiae) from the Great Barrier Reef. The Beagle, Records of the Northern Territory Museum of Arts and Sciencses 7: 65-78.
KOCH, G VON. 1878. Notiz über die Zooide von Pennatula. Zoologischer Anzeiger 1: 103-104.
KÖLLIKER, RA VON. 1870-72. Anatomisch-Systematische Beschreibung der Alcyonariarien. Erste Abtheilung: Die Pennatuliden. Abhandlungen von der Senckengbergischen naturforschenden Gesellschaft 7: 11-255; 487-602; 8: 85-275 (Also issued with consecutive pagination 1-458, dated 1872).
KÖLLIKER, RA VON. 1880. Report on the Pennatulida dredged by HMS Challenger during the years 1873-1876. Report of the scientific results of the voyage of HMS Challenger during the years 1873-76, Zoology 1: 1-41.
KÜKENTHAL, W. 1912. Der Stammbaum der Seefedern. Verhandlungen des Internationalen Zoologen-Kongresses, Jena 8: 563-570.
KÜKENTHAL, W. 1915. Pennatularia. Das Tierreich 43: 1-132. Verlag von R. Friedländer und Sohn, Berlin.
KÜKENTHAL, W. & BROCH, H. 1911. Pennatulacea. Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer "Validivia" 1898-1899 13: 113-576.
MARSHALL, A.M. 1887. Report on the Pennatulida dredged by HMS "Triton". Transactions of the Royal Society of Edinburgh 32: 119-152.
NIEDERMEYER, A. 1913. Über einige histologische Befunde an Veretillum cynomorium. Zoologischer Anzeiger 43: 263-270.
RETALLACK, G.J. 1994. Were the Ediacaran fossils lichens? Paleobiology 20(4): 523-544.
SCHMIDT, H. 1972. Die Nesseldapseln der Anthozoa und ihre Bedeutung für die phologentische Systematik. Helgoländer wissenschaftliche Meersuntersuchungen 23: 422-458.
SCHMIDT, H. 1974. On the evolution of the Anthozoa. Proceedings of the Second International Coral Reef Symposium 1: 533-560.
SEILACHER, A. 1989. Vendozoa: organismic construction in the Prterozoic biosphere. Lethaia 22(3): 229-239.
SOLÉ-CAVA, A.M. & THORPE, J.P. 1991. High levels of genetic variation in natural populations of marine lower invertebrates. Biological Journal of the Linnean Society 44: 65-80.
VERSLUYS, J. 1906. Bathyalcyon robustun nov. gen. nov. spec. Ein neuer Alcyonarier der Siboga-Sammlung. Zoologischer Anzeiger 30(17/18): 549-553.
WEERDT, W.H. DE. 1985. A sytematic revision of the North Eastern Atlantic shallow-water Haploscerida (Porifera, Demospongiae), Part I: Introduction, Oceanapiidae and Petrosiidae. Beaufortia 35(5): 61-91.
WEINBERG, S. 1976. Revision of the common Octocorallia of the Mediterranean circalittoral. I. Gorgonacea. Beaufortia 24 (313): 63-104.
WILLIAMS, G.C. 1989. The pennatulacean genus Cavernularia Valenciennes (Octocorallia: Veretillidae). Zoological Journal of the Linnean Society 95: 285-310.
WILLIAMS, G.C. 1990a. The Pennatulacea of southern Africa (Coelenterata, Anthozoa). Annals of the South African Museum 99: 31-119.
WILLIAMS, G.C. 1990b. A new genus of dimorphic soft coral from the southwestern fringe of the Indo-Pacific (Octocorallia: Alcyoniidae). Journal of Zoology 221(1): 21-35.
WILLIAMS, G.C. 1994. Biotic diversity, biogeography and phylogeny of pennatulacean octocorals associated with coral reefs in the Indo-Pacific. Proceedings of the Seventh International Coral Reef Symposium, 1992, 2: 729-735.
WILLIAMS, G.C. 1995a. Living genera of sea pens (Coelenterata: Octocorallia: Pennatulacea) - illustrated key and synopses. Zoological Journal of the Linnean Society (in proof).
WILLIAMS, G.C. 1995b. Revision of the pennatulacean genus Sarcoptilus (Coelenterata: Octocorallia), with descriptions of three new species from southern Australia. Records of the South Australian Museum 28(1): 15-34 (in proof).
WILLIAMS, G.C. 1995c. The enigmataic sea pen genus Gyrophyllum - redescription and phylogenetic assessment of S. sibogae from Tasmanian waters (Coelenterata: Octocorallia). Proceedings of the California Academy of Sciences 48(15) (in press).
{kind=link}